|
|
СИЛА МЕДЛЕННЫХ МЫШЕЧНЫХ ВОЛОКОН КАК ОСНОВНОЙ ФАКТОР ЛОКАЛЬНОЙ ВЫНОСЛИВОСТИ В
ЦИКЛИЧЕСКИХ ВИДАХ СПОРТА
Е.Б. Мякинченко, ПНИЛ
Введение. Имеется достаточное
число экспериментальных фактов, позволяющих считать, что большая мощность
энергетических и сократительных систем, локализованных непосредственно в мышцах
и определяющих т.н. локальную выносливость (ЛВ), позволяет отдалить наступление
утомления как сама по себе, так и путем снижения нагрузки на "центральные
факторы", интенсивное функционирование которых также может приводить к
утомлению и, следовательно, ограничивать спортивный результат в циклических
видах спорта (ЦВС). Этим определяется важность всех аспектов, касающихся
воспитания ЛВ в ЦВС.
На всех дистанция кроме спринтерских
(длительность - до 40 секунд) фактором спортивного результата, является
аэробная мощность мышц. С ним, при прочих равных условиях, связаны емкость
анаэробного гликолиза (это важно для средних дистанций) и аэробная
производительность (вклад в общее количество ресинтезируемой АТФ быстро
возрастает с увеличением дистанции). Считается, что аэробная мощность зависит
от массы ферментов митохондриальной системы. Наибольшей плотностью митохондрий
обладают медленные мышечные волокна (ММВ) и быстрые окислительные МВ (БоМВ).
Однако рекрутирование БоМВ и, тем более быстрых гликолитических МВ (БгМВ),
приводит к накоплению лактата, снижению рН мышц, быстрому расходу гликогена и
т.п., являющихся одной из причин утомления. Поэтому идеальным путем повышения
аэробной мощности мышц могло бы быть увеличение производительности ММВ. Как
этого добиться?
Основная часть. Считается, что аэробную
мощность повышает увеличение плотности митохондрий. Это действительно так,
однако в отношении БоМВ или БгМВ, но не в отношении ММВ. Это утверждение можно
обосновать следующим образом. При мышечной работе с мощностью аэробного порога
или ниже, когда в работу вовлечены, в основном, ММВ [Vollestad, 1985] продукция
лактата не превышает возможностей клеток по его утилизации. При этом нет
оснований сомневаться, что хотя бы часть медленных двигательных единиц работают
в условиях гладкого тетануса, т.е. - с максимальной мощностью. Это означает,
что мощности митохондриальной системы в этих МВ хватает, для ресинтеза
необходимого количества АТФ, расщепляемого на миофиламентах, в СПР и других
органеллах МВ. Другими словами, в ММВ мощность миозиновой АТФ-азы
сбалансирована с мощностью дыхательного фосфорилирования. В противном
случае, часть пирувата интенсивно восстанавливалась бы до лактата.
Следовательно, в процессе физической тренировки (по крайней мере у
квалифицированных спортсменов в ЦВС) отсутствует основной стимул для индукции
синтеза митохондриальных белков - дефицит энергии. Это подтверждается хорошо
установленным фактом, что низкоинтенсивная аэробная тренировка (не выше уровня
аэробного порога) обладает низкой эффективностью в отношении основных
показателей аэробной мощности.
Тогда закономерно возникает вопрос - какими
путями можно создать дефицит энергии в ММВ и, следовательно, стимулировать
повышение их аэробной мощности?
Традиционно для этого используется два подхода:
1) повышение мощности тренировочной работы и 2) использование искусственного
или естественного понижение напряжения кислорода во вдыхаемом воздухе, т.е. -
создание гипоксии мышц. В первом случае аэробная мощность действительно
повышается, однако за счет увеличения плотности митохондрий, капилляров в БМВ и
сопутствующей перестройки механизмов регулирования регионального кровотока. А
производительность ММВ, в общем случае, не растет (хотя есть исключения из этого
правила, о которых будет упомянуто ниже). Второй путь, теоретически, должен
быть более эффективен, но давать только временный эффект, так как, после
возвращения в условия нормобарии стимул исчезнет и, в соответствии с принципом
симорфоза, когда клетки избавляются от излишних структур, дыхательные
способности ММВ вернутся к исходу, несмотря на продолжающиеся напряженные
тренировки.
Таким образом, если в приведенных выше
рассуждениях нет принципиальных ошибок, то стратегическими путями повышения
аэробной мощности ММВ, гипотетически могут быть только два: (1) увеличение
АТФ-азной активности миозина и/или (2) увеличение числа миофибрилл. Обе эти
перестройки могут повысить энергозапрос во время тетанических сокращений и,
следовательно, создать стимул для увеличения массы всех ферментативных систем
энергообеспечения (креатинфосфокиназной и миокиназной реакций, аэробного и
анаэробного гликолиза, b -окисления жиров и др.). Первая возможность очень
гипотетична, так как АТФ-азная активность миозина зависит от типа иннервации
волокон, генетически обусловлена и, как считается, перестройке поддается с
большим трудом. Второй путь совершенно реален. Установлено, что при силовой
тренировке сила растет за счет накопления миофиламентов во всех типах мышечных
волокон.
В связи с этим возникают вопросы: 1) Не приведет
ли гипертрофия ММВ к снижению их аэробной мощности (ведь известно, что
представители силовых и скоростно-силовых видов спорта имеют низкие аэробные
показатели, а высокая степень и длительность ацидоза мышц, при силовой
тренировке могут иметь деструктивный эффект в отношении митохондриальной
системы)? 2) Реализуется ли путь гипертрофии ММВ у представителей ЦВС?
В дискуссиях по первому вопросу обычно
обсуждается модель Крога, предложенная в 1918 году, в которой участок ткани,
снабжаемый одним капилляром, рассматривается как цилиндр, осью которого служит
этот капилляр. Если принять допущение, что кислород внутри ткани перемещается
исключительно за счет свободной диффузии, то расстояние между капиллярами
становится критическим для адекватного снабжения всех точек клетки кислородом.
Это может привести к тканевой гипоксии. В качестве доказательства приводится,
например, то, что ткани, потребляющие в активном состоянии большое количество
кислорода, обладают наиболее развитой капиллярной сетью (миокард, красные мышцы
и др.); при аэробной тренировке окислительный потенциал мышц и капилляризация
волокон возрастают, как правило параллельно. Причем увеличение плотности
капилляров иногда происходит за счет уменьшения площади поперечного сечения
мышечных волокон, что интерпретируется как адаптивная реакция, направленная на
улучшение снабжения мышечной ткани кислородом за счет снижения диффузионного
расстояния [Б.С.Шенкман, 1990а, Т.Л.Немировская, 1992].
Однако существуют хорошо известные факты,
которые заставляют усомниться в бесспорности приведенных выше суждений.
1. В модели Крога не принимается во внимание
роль оксимиоглобина. Известно, что оксимиоглобин выполняет две функции: (1)
поддержание низкого внутриклеточного напряжения О2 для обеспечения высокого
градиента по отношению к капиллярной крови и (2) транспорт кислорода внутри МВ
в случае возникновения внутриклеточных градиентов кислорода. Поэтому наличие
оксимиоглобина уменьшает градиент парциального напряжение кислорода в разных участках
мышечных волокон, ликвидируя "гипоксические" участки.
2. Вокруг гипертрофированных оксидативных
мышечных волокон капиллярная сеть настолько густая, что средняя плотность
капилляров оказывается не менее высокой, чем в других местах мышцы [Gayeski, 1986].
Среднее межкапиллярное расстояние, при этом оказывается существенно меньше 80 m
м, которое считается критическим для адекватного снабжения ткани кислородом
даже на основании расчетов по модели Крога (т.е. - без учета роли
оксимиоглобина).
3. Прямые измерения показали, что напряжение
кислорода внутри мышечных волокон в состоянии максимальной респирации
митохондрий не зависит от размера волокон [Gayeski, 1986].
4. Показано, что в гипертрофированных мышечных
волокнах митохондрии располагаются по периметру волокна [Londraville, 1990].
Это уменьшает диффузионное расстояние и не вызывает необходимости накопления
миоглобина.
(1) |
(2) |
(3) |
(4) |
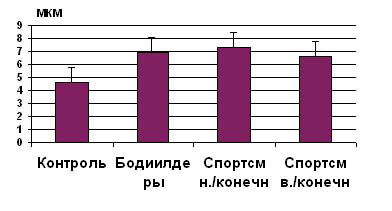
|
Рис.
1. Величина средней площади поперечного сечения (мкм2) по литературным данным [см.
обзор Е.Б.Мякинченко, 1997] медленных мышечных волокон у мужчин: (1) контроля, (2) элитных
бодибилдеров; (3) спортсменов тренирующихся на выносливость в мышцах нижних (бег, коньки, гребля,
велоспорт) и (4) верхних (плавание, гребля) конечностей.
5.
Существуют наблюдения [обзор Т.Л. Немировской, 1992]
о реципрокных отношениях между концентрацией миоглобина с одной стороны, плотностью
капилляров и окислительным потенциалом МВ - с другой в процессе объемной аэробной тренировки.
Следовательно, можно предположить, что система внутриклеточного транспорта кислорода обладает
резервами производительности, делающими ненужной высокую концентрацию миоглобина при
высокой капилляризации мышц (т.е. - при маленьком диффузионном расстоянии).
6. Существуют
многочисленные наблюдения о высокой степени гипертрофии мышечных волокон у спортсменов
экстракласса, тренирующих выносливость (рис. 1).
7. Аэробная
мощность и результат в ЦВС растет параллельно с гипертрофией ММВ. [Б.С.Шенкман, 1990б].
Однако
гипертрофия МВ бывает саркоплазматического или миофибриллярного типа [Г.Хопеллер, 1987]. Какой тип
реализуется у спортсменов в ЦВС? Известно, что объемная плотность митохондрий может возрасти, в
среднем, на 3-5% [Г.Хопеллер, 1987], количество гликогена - на 1%, гипотеза, что гипертрофия ММВ
может быть связана с большим количеством воды, связанной с гликогеном, не имеет
экспериментального подтверждения. Таким образом 50-100% гипертрофия ММВ может быть обусловлена, в
значительной мере, ростом массы миофиламентов и связанных с ними органелл клеток в результате
силовой тренировки. Меньшие показатели силы и скоростно-силовых способностей спортсменов,
тренирующих выносливость, [Hakkinen, 1988], могут быть связаны с меньшей степенью развития
"нервного" компонента мышечной силы.
Выводы. Приведенные
выше рассуждения позволяют выдвинуть парадоксальную, на первый взгляд, гипотезу, что
стратегическим путем повышения аэробных способностей мышц является гипертрофия ММВ
миофибриллярного типа. Тогда способом реализации этого пути будет оптимальное
сочетание аэробной тренировки с силовой, в которой основное воздействие осуществляется на
ММВ.
Литература.
1. Мякинченко Е.Б. Локальная выносливость в беге. М.: ФОН, 1997.-312с.
2. Hемировская Т.Л. Влияние аэробной тренировки на систему
доставки кислорода и энергетический метаболизм мышц человека. М.: Автореф. дисc. .... канд. биол.
наук., 1992, 23с.
3. Хоппелер Г. Ультраструктурные изменения в скелетной мышце под воздействием физической
нагрузки. М.: ЦООHТИ. ФиС. Выпуск 6., 1987,с. 3-48.
4. Шенкман Б.С. Влияние тренировки на композицию мышц, размеры и
окислительный потенциал мышечных волокон у человека. М.: //Автореф. дисс. ... канд. биол. наук, 1990.
23с.
5. Шенкман Б.С., Ширковец Е.А., Кузнецов С.Л. и др. Размеры и
окислительный потенциал мышечных волокон у гребцов. Hаучно-спортивный вестник, 1990, № 6, с.33-35
6. Gayeski T.E.J., C.R.Honig O2 gradients from sarcolemma to cell interior in red muscle at maximal VO2
//Am.J.Physiol. 251 (Heart Circ. Physiol.20):, 1986. H789-H799
7. Hakkinen K.,
K.L.Keskinen Muscle cross-sectional area and volantory force production characteristics in
elite strength- and endurance-trained athletes and sprinters //Eur. J. Appl. Physiol,
1989.59. 215-220.
8. Londraville R.L., Siddel
B.D. Maximal diffusion distance witthin skeletal muscle can be estimated from the
motochondrial distribution //Resp. Physiol, 1990. 3. 291-301
9. Vollestad NK, Blom PCS
Effect of varying exercise intensity on glycogen depletion in human muscle fibres. //Acta
Physiol Scand, 1985.125. 395-405
http://lib.sportedu.ru/Texts.idc?DocID=99215
|

